
Blackfly Biology
To see the illustrations full size, click on the image. To return to the text, use the BACK button on the Tool Bar
Blackflies are small flying insects with two wings belonging to the order Diptera, sub order Simuliidae. They are small, dark, stout-bodied and hump-backed (Fig.1) but they are generally larger than blood-sucking ceratopogonids (sandflies or midges) and range in size from 3mm to 7mm. They are also referred to as Simuliids. Wherever they cause a nuisance they usually have a special name in the local language.
| Description | Life Cycle | Pupa | Mating | Breeding sites | ||||
| Classification | Eggs | Adult | Feeding | Biting habits | ||||
| The Simuliidae | Larva | Habits | Oviposition | Response to varying conditions |
Head. Vision plays an important
role in the behaviour of blackflies. In the female the individual
elements (ommatidia) of which the eyes are composed are small
(10-15 µm) and the eyes are well separated above the antennae,
i.e. the female is dichoptic (Fig. 2). In the male the eyes are
much larger and touch above the antennae (holoptic), and the
lower  ommatidia are similar to those of the
female but the upper ones are greatly enlarged, measuring 25-40
1µm. The short horn-like antennae are the same in both sexes and
consist of small, disc-like segments, compacted together to give
a beaded appearance (Fig. 2). The commonest number of antennal
segments is 11; occasionally there are 10 as in the Australian (Austrosimulium)
species and rarely there are 9 in some North American Prosimulium
species. The 5-segmented, palps are considerably longer than the
short proboscis and carry on the third segment a large sensory
pit. In males and in a few species in which the females do not
bite, the mandibles and maxillae are not toothed.
ommatidia are similar to those of the
female but the upper ones are greatly enlarged, measuring 25-40
1µm. The short horn-like antennae are the same in both sexes and
consist of small, disc-like segments, compacted together to give
a beaded appearance (Fig. 2). The commonest number of antennal
segments is 11; occasionally there are 10 as in the Australian (Austrosimulium)
species and rarely there are 9 in some North American Prosimulium
species. The 5-segmented, palps are considerably longer than the
short proboscis and carry on the third segment a large sensory
pit. In males and in a few species in which the females do not
bite, the mandibles and maxillae are not toothed. Males differ from females by a larger
hump to the thorax, enlarged reddish eyes, and a more slender
abdomen which is tipped by a pair of claspers which are usually
visible
Males differ from females by a larger
hump to the thorax, enlarged reddish eyes, and a more slender
abdomen which is tipped by a pair of claspers which are usually
visible
Thorax. The wings are short (1.5 to 6.0mm) broad, colourless and transparent, with a large anal lobe (Fig. 3). The venation is characteristic, with well-developed radial veins along the anterior margin of the wing and weaker median and cubital veins posteriorly. In spite of its weak appearance the wing is highly efficient and in still air simuliids are capable of flying many kilometers. The radial sector may be unbranched or have two branches. Between the median (M2) and the cubital (Cu1) veins there is a forked submedian fold.
Abdomen. Made up of 8 segments, the last 3 of which compose the genitalia, and are not obvious The male terminalia are compact and relatively inconspicuous. The female has a single subspherical spermatheca.
Kingdom: ANIMALIA - Phylum: - ARTHROPODA
Class: INSECTA - Order: DIPTERA - Suborder: EUDIPTERA (or in older usage NEMATOCERA having whip-like many segmented antennae)
Infraorder: CULICOMORPHA - Family: SIMULIIDAE - Subfamilies: PARASIMULIINAE and SIMULIINAE
Genera: Many, of which Simulium and Prosimulium are most common
There are about 1809 valid simuliid species (including 11 are extinct) arranged in 28 genera of which four, Simulium, Prosimulium, Cnephia and Austrosimulium, are of economic importance, all of which belong to the Subfamily SIMULIINAE. The Subfamily PARASIMULIINAE contains just 4 species from western North America. The largest genus is Simulium with nearly 1200 species arranged in 42 subgenera. Simulium occurs in all zoogeographical regions with the greatest number (410) reported from the Palaearctic region. Prosimulium with 110 species assigned to six subgenera is largely confined to the Holarctic region, as is the genus Cnephia. Austrosimulium is restricted to Australia and New Zealand. The other large genera are Gigantodax with 65 species in the Neotropical region and Metacnephia with 51 species in the Holarctic. The Simuliidae are rich in species complexes most of which are only distinguishable by cytotaxonomy (i.e.by examination of their chromosomes), the best example being that of the African S. damnosum.
In Simulium and Austrosimulium the radial sector on the wing is unbranched; the costa bears spiniform setae and hairs and the hind leg has a rounded lobe (calcipala) at the inner apex of the first tarsal segment and a dorsal groove (pedisulcus) near the base of the second tarsal segment. In Prosimulium the radial sector is branched (sometimes only slightly), the costa bears only hairs and there is neither calcipala nor pedisulcus on the hind leg. In adults, species identification is often difficult requiring the use of microscopical characters such as the structure of the male and female terminalia. However, many species can be identified with relative ease using the respiratory organ of the pupal stage, and/or characters of the larval head.
The length of the life cycle varies with the species and
environmental conditions. In temperate regions species may have
one generation a year, while continuous breeding occurs in
tropical species. The larval stage of S. damnosum can be
completed in as little as 6 days, and the life cycle from egg to
adult can be completed in less than 2 weeks. 
The eggs, larvae and pupae are only found in moving water.
Ranging, according to species, from wave movement on lake shores
to small trickles in vegetation choked forest streams to the lip
of the highest waterfalls in the largest rivers. Distribution is
world-wide, they are absent only from deserts and a few isolated
islands. 
Eggs
The eggs are commonly laid in batches of 200-300, in a range of
30 to 800, on objects in or near running water or directly onto
water or on the surface. Eggs are either dropped directly into
the water and sink to the bottom or are laid on emergent objects
close to the waterline, where they are either directly wetted by
water or are in the splash zone. Communal egg masses may be
formed by several females ovipositing in close proximity, and
there is some evidence that gravid females may be attracted to
eggs laid by the same species, probably du to the presence of a 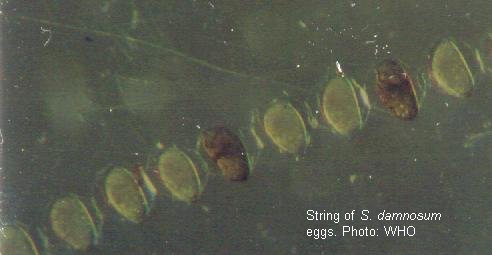 pheromone. Females of several species
crawl up to 15 cm below the water surface to oviposit on
submerged substrates.
pheromone. Females of several species
crawl up to 15 cm below the water surface to oviposit on
submerged substrates.
Eggs are 100 to 400 µm long and ovoid - triangular in shape. Their surface is comparatively smooth, lacking the patterned surface found in the eggs of Culicoides and culicids, and are covered with a gelatinous substance. Eggs may be laid in skeins like a string of beads, or in untidy masses. Freshly laid eggs are creamy-white, changing to dark brown or black within 24h. Simuliid eggs are sensitive to desiccation, even those of the Australian Austrosimulium. pestilens, which survive for many months in wet river deposits, desiccate rapidly when exposed to relative humidities of 96% or less. Eggs laid near the surface hatch when the embryo has completed development, a matter of days under favourable conditions. Other species produce dormant eggs in which the adverse conditions of summer and/or winter are passed.
Larva
The egg hatches to produce a larva which has a distinct,
sclerotized head with paired, simple eyes, and an elongated  hour-glass shaped body, in which the
thorax and posterior part of the abdomen are broader than the
anterior segments of the abdomen. The head bears a pair of
cephalic (labral) fans, homologous structures to the lateral
palatal brushes of mosquitoes. They do not create a current but
filter water passing over the larva. The larva has a single
anterior proleg, surmounted by a circlet of hooks and the abdomen
ends in a posterior circlet. The anus opens dorsally of the
posterior circlet, and from it may be extruded the rectal organ,
which probably, by analogy with the anal papillae of culicid
larvae, is concerned with chloride extraction from the water. The
larva spins a web of silk on the substrate, which is continued
into a silken thread on which the larva drifts downstream with
the current in search of a suitable object on which to settle.
When this has been found, the larva spins a patch of silk to
which it anchors itself by its posterior circlet of hooks. Larvae
normally remain near the surface of the water and are usually
found at depths of less than 300 mm (except for some large river
species which may be found at depths of several meters in
turbulent water). The larva can change its location by drifting
downstream on a silken thread, or by looping over the substrate
surface using the posterior circlet and the hooks on the anterior
proleg to retain a hold on secreted silk. Some species disperse
further from the oviposition site than others.
hour-glass shaped body, in which the
thorax and posterior part of the abdomen are broader than the
anterior segments of the abdomen. The head bears a pair of
cephalic (labral) fans, homologous structures to the lateral
palatal brushes of mosquitoes. They do not create a current but
filter water passing over the larva. The larva has a single
anterior proleg, surmounted by a circlet of hooks and the abdomen
ends in a posterior circlet. The anus opens dorsally of the
posterior circlet, and from it may be extruded the rectal organ,
which probably, by analogy with the anal papillae of culicid
larvae, is concerned with chloride extraction from the water. The
larva spins a web of silk on the substrate, which is continued
into a silken thread on which the larva drifts downstream with
the current in search of a suitable object on which to settle.
When this has been found, the larva spins a patch of silk to
which it anchors itself by its posterior circlet of hooks. Larvae
normally remain near the surface of the water and are usually
found at depths of less than 300 mm (except for some large river
species which may be found at depths of several meters in
turbulent water). The larva can change its location by drifting
downstream on a silken thread, or by looping over the substrate
surface using the posterior circlet and the hooks on the anterior
proleg to retain a hold on secreted silk. Some species disperse
further from the oviposition site than others.
In normal feeding position the larvae are anchored posteriorly
and extended in the direction of the current  with the head downsteam. The body is
twisted through 90-180° so that the fans and mouthparts face
towards the surface of the water. The water current is divided by
the proleg and directed towards the fans. A sticky secretion
produced by the cibarial glands enables the fans to capture fine
particles, which are transferred to the cibarium by the
mandibular brushes. Larvae of some species defend their territory
and are aggressive to their upstream neighbours, who would be
competing for the incoming food. Territorial defence declines
dramatically when food is abundant. Simuliid larvae ingest
particles such as bacteria, diatoms and silt up to 350 µm, but
the most commonly ingested particles are 10-100 µm. Algae pass
apparently unchanged through the blackfly gut but diatoms may
form as much as 50% of the gut contents. Filter-feeding larvae of
some species may also browse on the substrate while a few do not
filter feed at all but only graze on the substrate.
with the head downsteam. The body is
twisted through 90-180° so that the fans and mouthparts face
towards the surface of the water. The water current is divided by
the proleg and directed towards the fans. A sticky secretion
produced by the cibarial glands enables the fans to capture fine
particles, which are transferred to the cibarium by the
mandibular brushes. Larvae of some species defend their territory
and are aggressive to their upstream neighbours, who would be
competing for the incoming food. Territorial defence declines
dramatically when food is abundant. Simuliid larvae ingest
particles such as bacteria, diatoms and silt up to 350 µm, but
the most commonly ingested particles are 10-100 µm. Algae pass
apparently unchanged through the blackfly gut but diatoms may
form as much as 50% of the gut contents. Filter-feeding larvae of
some species may also browse on the substrate while a few do not
filter feed at all but only graze on the substrate.
Simuliid larvae are particularly abundant where the water
current accelerates, as at rapids, where presumably larvae will
strain a greater volume of water per unit time. Heavy larval
concentrations are often to be found at the outflows of large
lakes, where the water will be rich in phytoplankton for larval
food. Movement of water over the body surface provides the larva
with adequate dissolved oxygen for respiration. In deoxygenated
water larvae detach and drift downstream.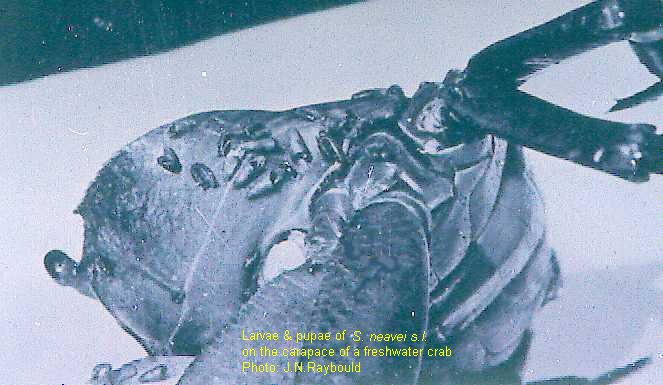 Simuliid
larvae reach a length of 4 to 12 mm, and being reasonably large
and aggregated, are easily seen on submerged objects. The larvae
of a few species are phoretic on freshwater crabs and mayfly
nymphs.
Simuliid
larvae reach a length of 4 to 12 mm, and being reasonably large
and aggregated, are easily seen on submerged objects. The larvae
of a few species are phoretic on freshwater crabs and mayfly
nymphs.
The mature last instar larva, recognised by the presence of a
dark "gill spot" through which the developing gills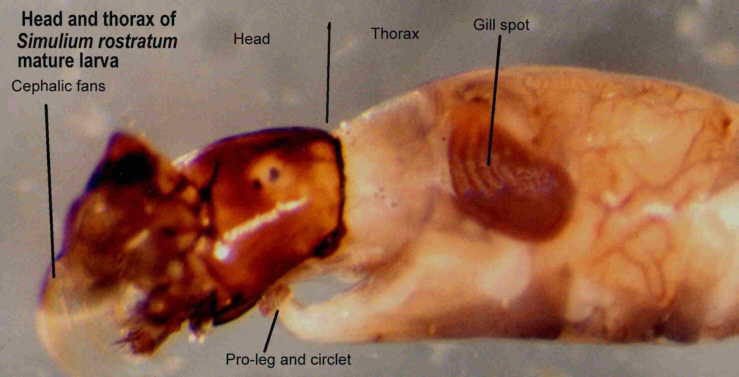 of the pupa may be seen, on the
lateral side of the thorax, is actually a pharate pupa within the
larval skin (i.e. midway between loosening the larval skin and
casting it off), and may move to a different site before
pupating. In most species the pharate pupa spins a cocoon in
which it pupates. This is usually slipper-shaped with the closed
end directed upstream and the open end downstream (Fig. 6). This
alignment prevents the cocoon being torn off the substrate by the
current. Construction of the cocoon takes about an hour and then
the larval skin is shed.
of the pupa may be seen, on the
lateral side of the thorax, is actually a pharate pupa within the
larval skin (i.e. midway between loosening the larval skin and
casting it off), and may move to a different site before
pupating. In most species the pharate pupa spins a cocoon in
which it pupates. This is usually slipper-shaped with the closed
end directed upstream and the open end downstream (Fig. 6). This
alignment prevents the cocoon being torn off the substrate by the
current. Construction of the cocoon takes about an hour and then
the larval skin is shed.
Pupa
The head and thorax of the pupa are combined into a single
cephalothorax, and there is a segmented abdomen (Fig. 6). The
latter bears spines and hooks which engage with the threads of
the cocoon and retain the pupa in place. The  cephalothorax bears a pair of
elongate, branched pupal gills, which trail downstream of the
cocoon. However, in some species the gills are short and barely
extend beyond the lip of the cocoon. The gills are homologous
with the respiratory horns of the Culicidae and Ceratopogonidae,
but they do not have open spiracles. The tubular branches of the
gill bear vertical struts which support a very thin, outer,
minutely perforated, trilaminate epicuticle and an inner fine
meshwork. The enclosed air-filled space around the struts
functions as a plastron. The shapes of the cocoon and gills are
important characters in the identification of species.
cephalothorax bears a pair of
elongate, branched pupal gills, which trail downstream of the
cocoon. However, in some species the gills are short and barely
extend beyond the lip of the cocoon. The gills are homologous
with the respiratory horns of the Culicidae and Ceratopogonidae,
but they do not have open spiracles. The tubular branches of the
gill bear vertical struts which support a very thin, outer,
minutely perforated, trilaminate epicuticle and an inner fine
meshwork. The enclosed air-filled space around the struts
functions as a plastron. The shapes of the cocoon and gills are
important characters in the identification of species.
The pupa, which does not feed, becomes progressively darker as the adult develops within. At emergence, the pupal skin splits, and the adult floats up to the surface in a bubble of air and immediately takes flight, or the newly emerged adult crawls up some emergent object to reach the air.
Adult
Adult emergence occurs predominantly in the daytime,
depending on light and temperature. In S. damnosum 60-90%
of the day's emergence has occurred by midday and there is no
emergence at night.
Very little is known about the activities of adult blackflies when not blood feeding or laying eggs, apart from the observation that both sexes may feed on plant sugars from extra-floral nectaries.
Mating
Mating occurs in the vecinity of the breeding site. In a few
species takes place on the ground but in the large majority of
species it occurs on the wing when males form small swarms in
association with visual markers which may be located 100 or 200 m
from the breeding site. Male simuliids recognize the female up to
a distance of 50 cm and pursue the female and attempt to couple.
There appears to be no contact pheromone because males will
attempt to mate with other males and with individuals of other
species. In some species male swarms and mating occur in close
proximity to the feeding sites of the females, which, for S.
ornatum and S. erythrocephalum, are respectively the
navel and ears of cattle. During mating a 2-chambered
spermatophore (capsule containing sperm) is transferred to the
female.
Feeding
Ornithophilic and mammalophilic species generally differ
morphologically in the shape of the claws on the last segment of
the legs. In mammal feeders the claws are usually simple but in
bird feeders they are toothed. This is assumed to be an
adaptation to holding and penetrating feathers but many species
feed indiscriminately on both avian and mammalian hosts.
Simuliids usually fly and feed out of doors, and are largely
diurnal. Some species will enter the natural openings of the
body, nose, ear and eye, behaviour which is particularly worrying
to livestock. As in other bloodsucking Nematocera both males and
females feed on nectar, which is stored in the crop. and only the
females are haematophagous with blood passing directly to the
midgut.
Female simuliids can be classified reproductively as showing obligate autogeny, i.e. females maturing all eggs without a blood meal; primiparous autogeny. i.e. females maturing the first egg batch without a blood meal but needing blood for each subsequent ovarian cycle; and more usually, obligate anautogeny when females need blood to mature every egg batch. When feeding, the female simuliid anchors its proboscis to the host by small hooks on the labrum and hypopharynx. The maxillae are protruded alternately, penetrating downwards and anchoring the proboscis more firmly. The mandibles cut into the skin with rapid scissor-like movements, penetrating to a depth of about 400æm. Blood is sucked upinto the stomach by the cibarial and pharyngeal pumps in the head. Blood-feeding takes about 4-5 min.
Oviposition
Oviposition habits vary greatly between species. S.
damnosum females oviposit communally in the short period
between tropical sunset and darkness. When flies are plentiful,
swarms of females lay their eggs on vegetation trailing in the
water,  returning to the same site every
evening, thus achieving densities of 2000-3000 eggs per square
centimetre. A. pestilens forms swarms of ovipositing
females which scatter their eggs over the surface of the water
where they become incorporated in the sandy river bed and can
survive two and a half years, providing they are kept permanently
damp. In Guatemala S. ochraceum drops its eggs directly
into the water; S. callidum lays its eggs one at a time on
the inclined surfaces of rocks; and S. metallicum, in
fast-flowing water lays its eggs on leaves without landing, and
in slower flowing water actually lands. Eggs of S. argyreatum
can withstand dryness during autumn and winter when the
temperatures are low, and they and the eggs of S. pictipes
resist frost and ice to survive the winter and hatch in the
spring.
returning to the same site every
evening, thus achieving densities of 2000-3000 eggs per square
centimetre. A. pestilens forms swarms of ovipositing
females which scatter their eggs over the surface of the water
where they become incorporated in the sandy river bed and can
survive two and a half years, providing they are kept permanently
damp. In Guatemala S. ochraceum drops its eggs directly
into the water; S. callidum lays its eggs one at a time on
the inclined surfaces of rocks; and S. metallicum, in
fast-flowing water lays its eggs on leaves without landing, and
in slower flowing water actually lands. Eggs of S. argyreatum
can withstand dryness during autumn and winter when the
temperatures are low, and they and the eggs of S. pictipes
resist frost and ice to survive the winter and hatch in the
spring.
Breeding
sites
Simuliids breed in running water, ranging from torrential
mountain streams to slow-moving lowland rivers, and a few species
are adapted to streams in which there is little perceptible
current. In Newfoundland it was found that the most significant
factors affecting the distribution of simuliid larvae were
current velocity, substrate type and water depth. A small number
of species have evolved a phoretic association with decapod
Crustacea (crabs, prawns) or Ephemeroptera (mayflies) in Africa
and the Himalayan region. Larvae and pupae of S. nyasalandicum
and S. woodi occur on the sides, the chelipeds, and the
basal segments of the walking legs of the crab Potamonautes
pseudoperlatus. They also occur on other species of
crab. Eggs are not laid on the crab and the young larva must find
its own phoretic partner. The most important of these phoretic
simuliid species is S. neavei, a vector of onchocerciasis.
In Africa mayfly phoretics are found mainly in heavily shaded,
forest streams and crab phoretics are found in small, forest
streams and larger and more open rivers.
When eggs are deposited in dense masses those in the outer layers hatch first and it is essential that the first instar larvae disperse. Larvae drift downstream attached to a silken thread or they can break the thread and drift with the current. Drifting and stationary larvae may form an important constituent of the diets of some fish.
Biting habits
Male simuliids do not require blood, and feed only on plant
sugars.
The females of most simuliids require a blood meal to develop
eggs. Some species feed on many birds and mammals including
humans and are probably an opportunistic feeders, whilst others
appear to be restricted to one or two host species. The
gonotrophic cycle may be remarkably short, being completed in 24
h in A. pestilens, but more usually in 3 to 4 days, with
eggs being laid very 5 or so days. 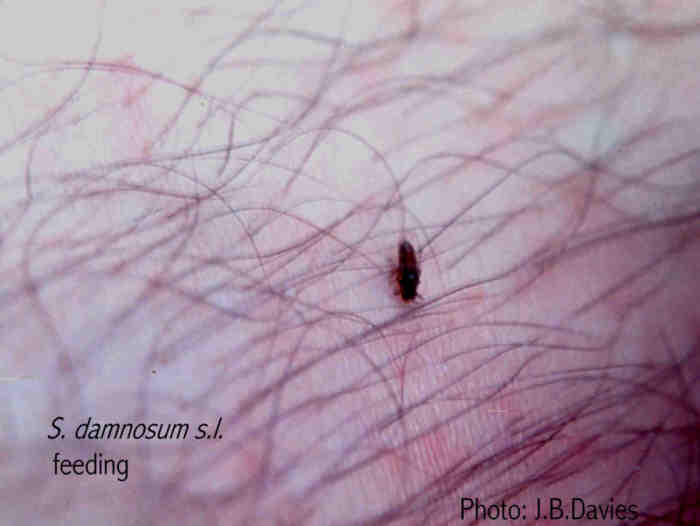
Simuliids are essentially diurnal insects, feeding in open sunny situations, but in the heat of the tropical midday activity may be suppressed leading to a bimodal pattern of activity with peaks around 0900 h in the morning and 1700 h in the afternoon. In shaded areas biting is more evenly distributed throughout the day. The circadian rhythm of biting activity varies with the age of the flies, with parous females feeding earlier in the day than nullipars. Dispersal has been most intensely studies in S. damnosum. There is linear dispersal along river courses in the gallery forest of the West African savanna; radial dispersal in the savanna during the rainy season, and in the forest region throughout the year; and differential dispersal, when nullipars disperse further than parous females, which tend to remain near the breeding sites.
There is also wind-borne dispersal. Simulium damnosum has been found to invade the Onchocerciasis Control Programme in West Africa from perennial forest breeding sites 200 to 400 km to the south-west, being carried on the early wet season SW monsoon winds. This explains their appearance in northern savanna country before the rivers begin to flow again after the dry season.
Response to varying conditions
Simuliids have to survive periods when the temperature
is too low to sustain normal activities, or when rivers cease to
flow in the dry season, which may be of indeterminate length.
Larvae of P. mixtum and P. fuscum grow actively
during the winter at temperatures near freezing point but larvae
of P. mysticum are dormant below 4°C reducing their
respiratory rates substantially.
In the severe climate of high latitudes simuliids show various adaptations to survival, involving reducing the time spent in the adult stage, which is the most vulnerable to low temperatures and high winds. Eight out of the nine species restricted to the Canadian tundra, e.g. S. baffinense, are autogenous and have reduced mouthparts; their females do not need to seek a blood meal and are unable to feed. The risk to the species' survival if the males had to swarm is avoided by mating occurring on the ground where adults cluster near the breeding site. As an adaptation to that behaviour the eyes of the male are sometimes dichoptic as in the female. As a further adaptation, a species may be both autogenous and parthenogenetic with the adult female becoming gravid in the pupa, e.g. P. ursinum, where there is virtually no free adult life.
Dry climates pose other problems. A. pestilens survives a dry season of uncertain length as viable eggs deep in the moist, sandy beds of transient rivers. Eggs of C. pecuarum laid in April remained dormant until November before developing and hatching in December. This carried the species over the summer months when many breeding places ceased to flow.
Many species of Simulium have several generations a year. When such species overwinter as growing larvae, as in S. monticola, there are likely to be large size differences between adults produced in the different seasons. Winter larvae produce larger adults than summer larvae. Changes in the total biovolume of the adult were found to be inversely related to mean water temperature